


")





Autor: Lucas Piroca. Vetanco Brasil.
La porcinocultura mundial continúa siendo presionada cada vez más para mantenerse rentable, al tiempo que tiene que atender la demanda de la población. Para llegar a los padrones actuales de desempeño y eficiencia productiva, la porcinocultura necesita de grandes avances en lo que refiere a tecnologías relacionadas a la nutrición, genética y sanidad, así como también en mejorías en las técnicas de manejo y de ambientación (GAVIOLLI et al., 2012).
Sin embargo, en base en los relatos de Cantarelli y Amaral (2013) la intensificación en la producción porcina generó un aumento de los desafíos dentro de las granjas y también, con el desenvolvimiento de estas tecnologías, aún permanecen algunos puntos críticos que limitan resultados óptimos para la productividad, llevando a perdidas económicas. Factores como la alta variabilidad y bajas de peso de los animales, así como la reducida ingestión de alimento y aumento de peso en algunos momentos continúan siendo los principales puntos críticos.
Estas preguntas nos llevan a pensar en el desenvolvimiento funcional de el tracto digestivo, el cual está relacionado al aumento de la salud intestinal que envuelve, entre otros factores, al desenvolvimiento y equilibrio anatómico/fisiológico de la microbiota y del sistema inmunológico (CANTARELLI; AMARAL, 2013).
El rendimiento de los animales depende de la integridad del tracto intestinal que debe permanecer saludable y funcional del inicio al fin de la vida, para reflejar en una óptima productividad. (GAVIOLLI et al., 2012).
EL INTESTINO DEL CERDO
El intestino del cerdo es, sin dudas, uno delos órganos que experimentó una extraordinaria transformación en un periodo de tiempo muy corto. Dentro de los mamíferos ocupa el segundo lugar, luego de la rata, en desarrollar dichas transformaciones. En menos de 50 días se multiplica alrededor de 4 veces transformando el tamaño de los valores iniciales de este órgano (SORACI et al., 2010).
El intestino, por diferencias estructurales, se divide en dos partes: Intestino Delgado e Intestino Grueso. El intestino delgado comprende el duodeno (4-4,5%), jejuno (88-91) y el Íleon (4-5). La proporción del duodeno en el neonato (recién nacido) es semejante al del adulto, mientras que la diferencia entre el jejuno y el Íleon no es clara, si bien existen características comunes en relación al intestino grueso, porción sin vellosidades formada por el cecon, colon y recto. El cerdo tiene un cecon relativamente corto y un colon largo. El cecon es un saco ciego cilíndrico localizado en la extremidad próxima al colon. El cecon, o colon ascendente, es transverso y la porción proximal del colon descendiente, están dispuestos en una serie de espirales centrífugos y centrípeta conocidas como colon espiral. El recto es incorporado en la gordura y es dilatado para formar una ampolla inmediatamente antes de terminar en el ano (LÆRKE & HEDEMANN, 2012).
El intestino de un cerdo adulto mide cerca de quince veces el tamaño de su cuerpo (aproximadamente 20 metros el intestino delgado y de 7,5 metros el intestino grueso) y tiene como función la digestión y absorción de nutrientes esenciales para los procesos metabólicos de los animales (ARAUJO; TEIXEIRA;VALENTE, 2010).
Según Soraci et al. (2010), durante mucho tiempo se pensó que digestión y absorción eran tareas a las cuales solamente se limitaba el intestino. Numerosos trabajos de investigación han demostrado la contribución del intestino al plano metabólico e inmunológico general del animal. En consecuencia, suministrar un alimento de excelente calidad, balanceado en su composición para una categoría de animal determinada, no garantiza el buen desarrollo de parámetros zootécnicos. El tubo digestivo debe encontrarse en condiciones fisiológicas óptimas para metabolizar los nutrientes aportados, como así también aportar la continua e importante entrada de antígenos orales. El correcto equilibrio de las funciones intestinales representan la clave para la obtención de una buena“performa”productiva.
MUCOSA INTESTINAL
La mucosa intestinal es una estructura que participa en los procesos de digestión –absorción de nutrientes- y provee una barrera fisicoquímica, metabólica e inmunológica contra la entrada de compuestos xenobíticos y toxinas, macromoléculas y microorganismos (bacterias, virus y hongos) en el organismo. La mucosa intestinal del cerdo tiene una superficie aproximada de 300 m², superficie equivalente a una cancha de tenis. (SORACI et al., 2010).
La mucosa del intestino delgado presenta varias estructuras que aumentan su superficie, incrementando el área disponible para absorción de nutrientes. El revestimiento del intestino delgado presenta una serie de pliegues permanentes, en forma circular, que consisten en dobras de la mucosa y submucosa. Ya las vellosidades intestinales son proyecciones alargadas de la mucosa con cerca de 0,5 – 1,5 μm de longitud, en dirección al lumen del intestino delgado. Entre las vellosidades existen pequeñas aberturas de glándulas tubulares simples denominadas Criptas (JUNQUEIRA;CARNEIRO,2011).
Las vellosidades intestinales, además de contribuir para la defensa contra una infección, también tienen como función aumentar la superficie de absorción de los nutrientes previamente digeridos, siendo así asociados como un importante parámetro morfológico de validación de desenvolvimiento animal (PLUSKE, HAMPSON;WILLIANS, 1997).
Las vellosidades y las Criptas son recubiertas por una capa continua de epitelio celular intestinal (enterocitos) que se disponen en forma de monoestrato y en relación con células endocrinas, inmunes y células globulares productoras de mucus, como ilustra la Imagen 1. La morfología epitelial cambia a lo largo del tracto intestinal, pero en esencia consiste en una región de criptas (donde se encuentran las células madres y de Paneth, que participan en funciones de defensa) y una región apical (vellosidad) donde las células se encuentran en diferente grado de diferenciación y función.
Las células epiteliales que alcanzan un determinado grado de diferenciación y maduración, mueren y son liberadas all lumen del intestino . De hecho, las células de las Criptas intestinales están entre las células de los organismos que se regeneran más rápidamente, representando la mayor necesidad de la síntesis de proteínas aisladas de animales que no están en crecimiento. La velocidad de recambio de las células del epitelio intestinal del cerdo es muy rápida, oscilando entre 2 a 5 días (SORACI et al., 2010).
Imagen 1. Estructura anatómica del epitelio intestinal y células que la componen
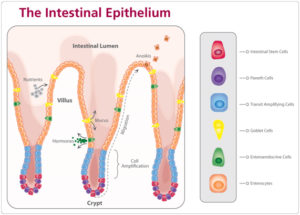
Cuando las células de las criptas se multiplican, ellas migran para la base de la vellosidad, empujando otras células de la vellosidad para frente de ellas, de modo que hay una progresión continua de células que migran para las vellosidades. Conforme migran, las células se tornan maduras y pasan de células relativamente indiferenciadas en las criptas a células altamente absorsivas especializadas en las vellosidades. El largo de las vellosidades es determinado por la velocidad en la cual las células perdidas en el ápice de las vellosidades son sustituidas por las células de las criptas (HERDT, 2014).
El aumento de la productividad de la Cripta está asociada con el incremento del número de células secretorias, que pueden influenciar en la patogénesis de la diarrea (PIERCE ET., 2005). Sin embargo, el aumento de la profundidad de la cripta puede ser debido a la mayor producción de ácidos grasos, estimulando la multiplicación celular en el intestino delgado (MONTAGNE;PLUSKE; HAMPSON, 2003).
MUCUS INTESTINAL
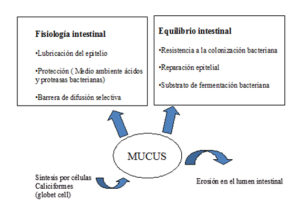
Soraci et al, (2010) definen el mucus intestinal como una biocapa de cobertura de la mucosa producida por secreción de estas células caliciformes. El mucus constituye una berrera de defensa contra microorganismos y agentes físicos-químicos. A su vez, cumple funciones de lubricación y transporte entre el contenido presente en el lumen intestinal y la superficie del epitelio. Desde el punto de vista químico, se trata de un gel compuesto por un 95% de agua y electrólitos, carbohidratos, aminoácidos, proteínas y lípidos.
Sus propiedades bioelásticas responden a sub unidades de mucina, proteína unida a largas cadenas de carbohidratos que contienen un azúcar neutral de hexosamina. Los carbohidratos poseen en sus cadenas terminales grupos de sulfatos libres unidos a ácido siálico.
Uno de los mecanismos de defensa del mucus contra microorganismos patogénicos, se relaciona con la capacidad que tiene la mucina de establecer uniones a través de sus carbohidratos terminales con receptores en la superficie de determinadas bacterias. Dicha interacción impide la fijación a las células epiteliales y su posterior destrucción o invasión intracelular (SORACI et al., 2010).
MICROBIOTA
Además de los factores maternos, del destete y de la edad, con mucho énfasis ha sido colocado el papel de la microbiota en el desenvolvimiento de la función intestinal e inmunidad. Estudios en lechones sin gérmenes han destacado la importancia de una microbiota intestinal para el desenvolvimiento del sistema inmune de la mucosa intestinal (STOKES et al., 2004).
El tracto gastrointestinal del cerdo contiene 10¹⁴ microorganismos procariotas y eucariotas, valor que resulta 10 veces más alto que el número de las células del cuerpo. La microbiota de un cerdo alberga más de 400 especies de bacterias. Esos microorganismos que colonizan el intestino se distribuyen en comunidades extremamente diferentes y ejercen un marcado efecto en la filosofía intestinal, a nivel morfológico, secreción de mucus, digestión de nutrientes, metabolismo y función inmune. Aunque la microbiota intestinal representa un sistema complejo de interacciones, el rol más estudiado tiene que ver con el efecto protector de determinadas especies de bacterias: Lactobacilos y Bifidobacterias contra infecciones entéricas. El mecanismo involucrado se relaciona con una interacción simbiótica con el epitelio intestinal y el sistema mucosal inmune (SORACI et al., 2010).
La microbiota presente en el aparato digestivo puede consultar protecciones físicas, químicas y biológicas (ZHU et al., 2002). Mientras tanto, el desempeño de los animales puede ser perjudicado debido a la producción de sustancias tóxicas generadas por la actividad de determinadas bacterias patogénicas, también presentes en la microbiota intestinal, lo que determina el espesor de la pared intestinal y causa la menor utilización de los nutrientes (WALTON, 1990; PEDROSO; OETTING; UTIYAMA, 2005). Siendo así cabe a nosotros conocer la microbiota, qué y cómo podemos hacer para buscar el equilibrio de la misma.
Funciones de la microbiota según Soraci et al. (2010)
1. Participar en la mantención del equilibrio e integración del epitelio intestinal;
2. Barrera protectora competitiva y química (bacteriocitas, fermentación ácida que mantiene un pH relativamente bajo) frente a la invasión de microorganismos;
3. Incrementar la absorción de nutrientes, particularmente mineral (Ca++);
4. Participar en la síntesis metabólica de vitaminas (grupo B y C) y de ácidos graso volátiles de cadena corta (acético, propiónico y butírico);
5. Estimular el desarrollo de respuesta inmune (A través de comunicaciones cruzadas o cross-talk o “negociaciones” entre la microbiota intestinal y el huésped); y
6. Metabolismo de urea, sales biliares y ácidos grasos.
INMUNIDAD INTESTINAL
El sistema inmune mucosal desempeña una doble función. Por una parte, debe identificar nutrientes inocuos y suprimir cualquier respuesta inmune sistémica que se pudiera generar contra ellos. Por otra parte, debe reaccionar para excluir cualquier invasión por virus, bacterias, parásitos y hongos (SORACI et al., 2010).
El lechón es profundamente inmunodeficiente al nacer y es altamente dependiente de una composición específica y no específica de factores inmunológicos presentes en el calostro y leche materna para protección inmunológica, desarrollo y supervivencia. Después del nacimiento, el lechón absorbe macromoléculas de forma no selectiva.
IgG del calostro de la cerda es absorbida por el recién nacido vía enterocitos (KOMUVES; NICOLS; HUTCHENS, 1993). Un “bloqueo” del intestino para la captación macromolecular ocurre dentro de 24 hs a 48 hs después del nacimiento según Leece (1973 apud STOKES et al., 2004). Los linfocitos inmunológicos necesarios para la producción de IgA secretoria que están en la leche de la cerda para mantención de la inmunidad humoral de la prole (SALMON, 2000).
Los cerdos recién nacidos solo son capaces de generar respuestas limitadas de células T y B cuando son enfrentados con patógenos, caracterizando el estado de inmunocomprometido. (BUTLER, 2002). Claramente, el desenvolvimiento de la inmune competencia es un requisito absoluto para el óptimo crecimiento y desempeño.
Sin embargo, en el contexto de la exposición a una vasta gama de antígenos asociados a patógenos y con bacterias comensales y alimentares, una definición de inmune competencia debe considerar la posibilidad de montar respuestas a los antígenos, eso incluye la capacidad de generar tolerancia a los alimentos y antígenos bacterianos comensales, más allá de las respuestas inmunes activas a patógenos (BAYLEI et al., 2001).
Según Stokes et al., (2004) existe un conjunto considerable de pruebas para indicar el papel ambiental de la nutrición en el desenvolvimiento inmunológico. Hay también una visión de que los sistemas inmunológicos requieren estimulación durante periodos muy definidos de desenvolvimientos post natal precoz, con la finalidad de que la función inmunológica óptima y la que falla en entregar esos insumos pudiendo perjudicar la función inmunológica en el inicio y en toda la vida. Dependiendo de la composición alimenticia y de la presencia de componentes alimenticios orales a la morfología, tanto del intestino como de la mucosa y las células inmunes intestinales sufren mudanzas.
Las células epiteliais intestinales son ahora reconocidas por desempeñar un papel importante en la inmunidad de forma innata, formando un cuerpo físico y barrera funcional a la dieta y antígenos microbianos. Estas células responden a bacterias colonizadas utilizando células de reconocimiento con padrones para detectar, como para responder a su presencia. Sistemas receptores diversos son expresados en las superficies de las células epiteliais que reconocen las bacterias y pueden comunicarse por diversas señales con las poblaciones de células linfoides.
Según Soraci et al. (2010), el sistema inmune intestinal del cerdo cuando es maduro depende de una inmunidad no especifica a cargo de células Killer mast cell, APC: Células presentadoras de antígenos, células T-helper, macrófagos y neutrófilos, que actúan a través de mecanismos quimiotácticos y de mecanismos de respuesta inmune a cargo del tejido asociado al intestino (Galt – gut associated lymphoid tissue,) El tejido linfoide representa un 30% de la masa intestinal y el 50% del tejido linfoide del organismo. Se estiman que en el cerdo existen alrededor de 10¹⁰ células productoras de anticuerpos¬/m del intestino. El mismo se encuentra a un nivel intestinal formando 2 partes:
1. Organizado en placas (Placas de Peyer) y nódulos linfáticos intestinales.
2. Células inmunes diseminadas en forma difusa a lo largo del tracto intestinal (lámina propia y células intra-epiteliales).
Las placas de Peyer están formadas por múltiples folículos (células B) rodeadas por zonas interfoliculares (Células T). En la lámina propia, las células plasmáticas (células B maduras) se encuentran situadas en las criptas, mientras que a las células T (CD4+ y CD8+) se las encuentran en la vellosidad.
Los receptores Toll-like (TLRs) están surgiendo como una clase funcionalmente importante de receptores de membrana con papeles fundamentales en el reconocimiento e inmuno modulación. TLRs reconocen padrones moleculares conservados (padrones moleculares asociados a patógenos – PAMPs y padrones moleculares asociados a comensales CAMPs) compartidos por grandes grupos de bacterias y otros microorganismos intestinales.
Estos mecanismos TLR desencadenan la liberación de muchas citosinas y quimiocinas que pueden ejercer una profunda influencia sobre las células del sistema inmune innato y adaptativo (STOKES et al. 2004).
El sistema inmune específico participa a través de una respuesta humoral (normalmente dirigida a bacterias ) y una respuesta celular (dirigida a células infectadas por virus).
INMUNOGLOBULINAS
IgA e IgM: Representan la barrera inmunológica secretoria principal en el cerdo (IgM, particularmente en animales jóvenes). La IgA es sintetizada por los linfocitos B de las placas de Peyer. La IgA brinda protección al prevenir la adherencia de bacterias y toxinas a las células epiteliales, proceso conocido como exclusión inmune.
IgE: Esta inmunoglobulina se encuentra asociada a las células cebadas y de la lámina propia. Su importancia radica en la protección contra infecciones parasitarias y en la regulación y ampliación de respuesta inmune local
La mantención de la integridad de la superficie epitelial es un proceso critico en la prevención de infecciones. No es también sorprendente que los mecanismos de defensa inmune de la mucosa tiendan a no ser inflamatorias y principalmente direccionadas para mantener los potencialmente antígenos dentro del lumen del intestino, donde el peristaltismo y el flujo constante de la digestión irán efectivamente a removerlos.
La integridad física (células epiteliais y producción de mucus) conjuntamente con IgA fueron una barrera a la entrada de substancies potenciales antigénicas. Solamente cuando la barrera es violada por otros procesos defensivos desempeñan un papel.
INMUNIDAD – PROTECCIÓN Y TOLERANCIA DEL INTESTINO MADURO
Según Stokes et. al. (2004) la indicación de respuestas inmunes adaptativas comienzan con el procesamiento y presentación de antígenos por estructuras específicas, probablemente en los tejidos organizados como placas de Peyers o linfonodos mesentéricos. Las células dendríticas presentan un antígeno clave y son importantes para el desenvolvimiento de la inmunidad innata y adaptativa. La respuesta subsiguiente a los antigenios de la mucosa, ya sea en busca de respuestas inmunes de anticuerpos celulares, IgA o todavía tolerancia inmunológica, depende de la exposición anterior a células dendríticas.
Moléculas de conexión a la TLR, pueden envolver el subsecuente reconocimiento del antígeno por células T primarias en microambientes de citosinas tales como la lámina propia intestinal. El resultado al antígeno es un paso crítico en la mantención de “la salud entérica”y requiere la capacidad de discriminar entre destruir patógenos y responder adecuadamente a antígenos inofensivos. Estudios realizados en la última década identifican varios mecanismos por los cuales las respuestas inmunes en el intestino son regulados. Estos incluyen la eliminación clonal, anergia clonal y control activo por células reguladoras. La anergia clonal es una característica de dosis muy elevadas de antígeno alimenticio por en cuanto la inducción de células T reguladoras (Treg) es importante en los niveles fisiológicos de exposición.
DESENVOLVIMIENTO DE LA MUCOSA Y ARQUITECTURA INMUNOLÓGICA
Stokes et al. (2004) afirma que al contrario del cerdo adulto, donde existe un elevado grado de organización, trabajos han demostrado que las células y estructuras involucradas en la respuesta inmune de la mucosa están inicialmente ausentes al nacer y hay una secuencia en la maduración intestinal del lechón.
La arquitectura del intestino es comparable a la del animal maduro solamente a la 7ª semana. Mientras la tolerancia a proteínas alimentares parece no estar totalmente pronta hasta la 8ª semana.
FACTORES DE RIESGO
En lo que se refiere a salud intestinal y producción de cerdos, encontramos diversos factores de riesgo, dentro de ellos micotoxinas, oxidación de lípidos, calidad de los alimentos, estrés y errores de manejo y ambientación los cuales generalmente llevan a enteritis infecciosas.
Las enteritis infecciosa por sí mismas, están entre los factores de riesgo que representan el mayor problema en la creación técnica de cerdos, debido a las grandes perdidas económicas asociadas con estas patologías. Los problemas recurrentes a diarreas en cerdos son consecuencia de costos con antibióticos, el aumento de la mortalidad, necesidad de manejo y cuidados con los animales enfermos y conversión alimenticia inadecuada (HOLLAND, 1990). El conocimiento de la patogenicidad de las diarreas en cerdos es sobre los patógenos responsables, es una importante herramienta para el diagnóstico de las enfermedades causadas por los mismos. De esta forma, medidas de tratamiento y control pueden ser tomadas minimizando las pérdidas económicas acarreadas de problemas entéricos (ZLOTOWSKI; DRIEMEIER; BARCELLOS, 2008).
En base a esto se destaca abajo en la tabla 1 los principales patógenos relacionados con diarreas en cerdos, conjuntamente a los mecanismos básicos de acción de estas diferentes localizaciones del tracto intestinal y aspectos generales del excremento.
Tabla 1. Localización, mecanismos de acción y aspecto de los excrementos infectados con los principales patógenos entéricos de los cerdos.
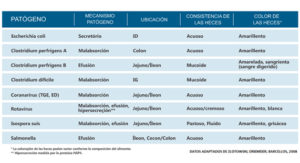
La correlación de las informaciones presentadas en las tablas 1, 2 y 3 en conjunto con una anamnesis bien conducida, estrechan los caminos para llegar a un diagnóstico presuntivo y posterior confirmación con análisis de laboratorio.
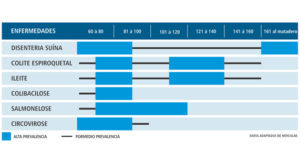
Tabla 2. Principales enfermedades en la fase de maternidad y vivero con su respectiva prevalencia por periodo semanal
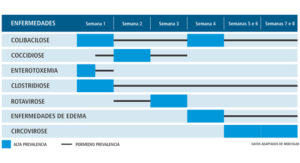
Tabla 3. Principales enfermedades de la fase de crecimiento y terminación con respectiva prevalencia por periodo
CONCLUSIONES
Un intestino saludable es aquel que presenta adecuada integridad anatómica conjuntamente a la mantención del equilibrio entre su inmunidad fisiológica y microbiota. Toda acción que tomamos, sea en el ámbito nutricional o sanitario, como también el ambiente que disponemos para el cerdo y la forma como lo manejamos, interfiere directamente en la salud intestinal.
Tenemos innumerables trabajos científicos demostrando el retorno financiero de las inversiones en ambientación, manejo, nutrición y sanidad en dirección a la salud intestinal. Por más que todavía téngamos mucho que descubrir y/o entender mejor sobre salud intestinal y los factores relacionados a la misma, ya tenemos establecidos cuáles son los principales agentes patógenos, qué enfermedades causan, cómo los diagnosticamos y principalmente lo que nos compete hacer.
Siendo así, nos cabe juntar el conocimiento científico que tenemos en nuestras manos y llevarlo a las granjas donde lo pondremos en acción visando en el bienestar de los animales y un mejor retorno financiero en la cadena de producción de cerdos.
REFERENCIAS
ARAUJO, A. C. P.;TEIXEIRA FILHO, A.; VALENTE, A. L. Anatomia dos animais de produção II. Pelotas: Universidade Federal de Pelotas, 2010. (Apostila).
BAILEY, M.; PLUNKETT, F. J.; ROTHKOTTER, H. J.; VEGA-LOPEZ, M. A.; HAVERSON, K.; Stokes, C.R. Regulation of mucosal immune responses in effector sites. Proc. Nutr. Soc. 60, 2001. p. 427-435.
BUTLER, J.E.; WEBER, P.; SINKORA, M.; BAKER, D.; SCHOENHERR, A.; MAYER, B.; FRANCIS, D. Antibody repertoire development in fetal and neonatal piglets. VIII. Colonization is required for newborn piglets to make serum antibodies to T-dependent and type 2 T-independent antigens. J. Immunol. 169, 2002. p. 6822–6830.
CANTARELLI, V. S.; AMARAL, L. G. M. Desenvolvimento do sistema digestório de leitões. In: 28º REUNIÃO ANUAL DO CBNA: CONGRESSO SOBRE NUTRIÇÃO DE ANIMAIS JOVENS – AVES E SUÍNOS, 2013, Campinas. Anais…Campinas: CBNA, 2013. p.1-13.
GAVIOLI, D. F.; OLIVEIRA, E. R.; SILVA, A. A.; ROMERO, N. C.; LOZANO, A. P.; SILVA, R. A. M.; BRIDI , A. M.; OBA, A.; SILVA, C. A. Efeito de promotores de crescimento para suínos sobre o desempenho zootécnico, a qualidade intestinal e a eficiência da biodigestão dos dejetos. Semina: Ciências Agrárias, Londrina, v. 34, n. 6, suplemento 2, 2013. p. 3983-3998.
HOLLAND, R. E. Some infectious causes of diarrhea in young farm animals. Clinical Microbiology Reviews. 3, 1990. p. 345-375.
HERDT, T. Digestão e absorção: os processos não fermentativos. In: CUNNINGHAM, J. G. Tratado de fisiologia veterinária. 3.ed. Rio de Janeiro: Guanabara Koogan, 2004. p. 263-287.
JUNQUEIRA, L. C.; CARNEIRO, J. O trato digestivo. In: ______. Histologia básica. 11. ed. Rio de Janeiro: Guanabara Koogan, 2011. p. 283-316.
KOMUVES, L. G.; NICOLS, B. L.; HUTCHENS, T. W.; HEATH J. P. Formation of crystalloid inclusions in the small intestine of neonatal pigs: an immunocytochemical study using colloidal gold. Histochem. J. 25, 1993. p. 19–29.
LÆRKE, H. N.; HEDEMANN, M. S. The digestive system of the pig.
Nutritional physiology of pigs – Online Publication. ed. / Knud Erik Bach Knudsen; Niels Jorgen Kjeldsen; Hanne Damgaard Poulsen; Bent Borg Jensen. Foulum: Videncenter for Svineproduktion, 2012. p. 5-7.
MONTAGNE, L.; PLUSKE, J. R.; HAMPSON, D. J. A review of interactions between dietary fibre and the intestinal mucosa, and their consequences on digestive health in young non-ruminant animals. Animal Feed Science and Technology, Amsterdam, v. 108, n. 1-4, ago. 2003. p. 95-117.
PEDROSO, A. A; OETTING, L. L.; UTIYAMA, C. E. et al. Variabilidade espacial da comunidade bacteriana intestinal de suínos suplementados com antibióticos ou extratos herbais. Revista Brasileira de Zootecnia, Viçosa, v. 34, n. 4, 2005. p. 1225-1233.
PIERCE, K. M.; SWEENEY, T.; BROPHY, P. O.; CALLAN, J. J.; McCARTHY, P.; O’DOHERTY, J. V. Dietary manipulation post weaning to improve piglet performance and gastro-intestinal health. Animal Science, Champaign, v. 81, 2005. p. 347-356.
PLUSKE, J. R.; HAMPSON, D. J.; WILLIAMS, I. H. Factors influencing the structure and function of the small intestine in The weaned pig: a review. Livestock Production Science, Amsterdam, v. 51, 1997. p. 215-236.
SALMON H., Mammary gland immunology and neonate protection in pigs – Homing of lymphocytes into the MG. Biology of the Mammary Gland 480, 2000. p. 279–286.
SINGH, B.; READ, S.; ASSEMAN, C.; MALMSTROM, V.; MOTTET, C.; STEPHENS L. A.; STEPANKOVA, R.; TLASKALOVA, H.; POWRIE, F. Control of intestinal inflammation by regulatory T cells. Immunol. Rev. 182, 2001. p. 190–200.
SORACI, A. L.; AMANTO, F.; HARKES, R.; PÉREZ, D. S.; MARTÍNEZ, G.; DIEGUEZ, S. N.; TAPIA, M. O. Uso estratégico de aditivos: Impacto sobre el equilibrio y salud gastrointestinal del lechón. Analecta Vet 2010; 30 (1): 42-53, 2010.
STOKES, C. R.; BAILEY, M.; HAVERSON, K.; HARRIS, C.; JONES, P.; INMAN, C.; PIÉ, S.; OSWALD, I. P., WILLIAMS, B. A., AKKERMANS, A. D.L., SOWA, E.; ROTHKÖTTER, H-J.; MILLER, B. G. Postnatal development of intestinal immune system in piglets: implications for the process of weaning. HEALTHYPIGUT Workshop 1 – Digestive physiology, microbiology and immunology basis for gut function and dysfunction in pigs; Anim. Res. 53, 2004. p. 325-334.
WALTON, J. R. Modo de acción de los promotores de crescimento. Industria Porcina, [s. l.], v. 10, n. 2, 1990. p. 6-11.
ZHU, X. Y.; ZHONG, T.; PANDYA, Y.; JOERGER, R. D. Analysis of microbiota from cecum of broiler chickens. Applied and Environmental Microbiology, Washington, v. 68, n. 1, 2002. p. 124-137.
ZLOTOWSKI, P.; DRIEMEIER, D.; BARCELLOS, D. E. S. N. Patogenia das diarréias dos suínos: modelos e exemplos. Acta Scientiae Veterinariae. 36(Supl 1), 2008. p. 81-86.